
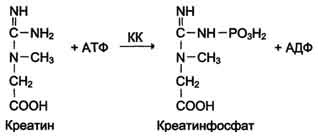
Креатинкиназа-АТФ: креатин-1,4-фосфотрансфераза (КФ 2.7.3.2) — катализирует обратимый перенос фосфатного остатка между АТФ и креатином с образованием АДФ и креатинфосфата. Общепринято сокращенное название фермента — креатинкиназа (КК). КК — цитозольный и митохондриальный фермент, который функционирует в клетках многих тканей. Продукт реакции фосфокреатин — макроэргическое соединение, обеспечивающее энергией сокращение мышцы, ее расслабление и транспорт метаболитов в миоцит. В первое время активность КК в крови интересовала клинических химиков как тест для выявления поражения поперечнополосатой мускулатуры. В настоящее время активность КК — ведущий тест в диагностике инфаркта миокарда.
Наибольшее содержание КК отмечено в миокарде и скелетных мышцах, высокое содержание — в языке, мышце диафрагмы, низкое — в почках, легких, печени. КК — димер; в цитозоле клеток тканей человека присутствуют 2 субъединицы: М — мышечный тип и В — мозговой тип. Субъединицы представляют собой белки с молекулярной массой 41кДа с активным центром в каждой субъединице. Соответственно двум формам субъединиц димерная форма молекулы КК имеет следующие варианты: мышечный тип КК-ММ, гибридный димер КК-МВ, характерный для миокарда, и изофермент КК-ВВ, локализованный преимущественно в мозговой ткани.
Активность фермента и распределение его изоферментов органоспецифичны, поэтому идентификация и количественное определение изоферментов КК в сыворотке крови не только полезно для установления увеличения проницаемости мембран или повреждения специфического органа, но и дает возможность установления диагноза, наблюдения клинического течения и оценки прогноза заболевания. В электрическом поле наибольшей подвижностью обладает КК-ВВ, наименьшей — КК-ММ, гибридная форма КК-МВ занимает промежуточное положение. В соответствии с этим в классификации Международного союза теоретической и прикладной химии КК-ВВ обозначена изоферментом I КК, КК-МВ — II, КК-ММ — III. Для понимания источников появления в крови активности КК-МВ следует отметить, что у плода до 16-недельного возраста в мышечной ткани доминирует зародышевая форма КК, а именно КК-МВ, которая позже замещается КК-ММ. Определенная перестройка изоферментного состава КК происходит и при гипертрофии миокарда: повышается процентное содержание КК-МВ и КК-ВВ, а доля КК-ММ оказывается сниженной. Миоциты в условиях длительной гиперфункции повышают синтез изоформы КК-ВВ.
Клиническое значение определения активности КК и ее изоферментов.
Повышение активности КК в крови может быть следствием травмы, переохлаждения или перегревания, голодания и бактериальной интоксикации, дегидратации, поражения электрическим током. Повышенная активность КК-МВ отмечена при мышечной дистрофии разной этиологии, ишемическом рабдомиолизе с миоглобинурией, отравлении угарным газом. Активность КК-МВ повышалась, более чем на порядок превышая норму, у больных острым перикардитом.
Опыт применения изоферментной диагностики в блоке интенсивной терапии показал, что повышение в крови активности КК-МВ может происходить при выраженной недостаточности кровообращения, катетеризации сердца, отеке легких, аритмиях, кардиогенном шоке, алкогольной интоксикации, кровотечении из ЖКТ, травматической эпилепсии, травме грудной клетки. Активность КК-МВ повышена в крови пациентов при синдроме шока, развившемся при анемии, гиперкапнии, гипоксемии, молочнокислом ацидозе, выраженной гипотонии. Однако только в редких случаях в указанных ситуациях активность КК-МВ на 6% превышала активность КК. Учитывая возможности неспецифического повышения активности КК-МВ, надо помнить и о возможности пропустить небольшой инфаркт миокарда. У больных с гипертрофией миокарда активность КК-МВ в период инфаркта может достигать 1/3 активности КК.
Повышение активности КК в сыворотке крови может быть обусловлено увеличением активности изофермента КК-ММ.
тивность КК-ММ в сыворотке крови повышается при прогрессирующей мышечной дистрофии, дерматомиозите, полимиозите, инфаркте миокарда, гипотиреозе, некоторых заболеваниях нервной системы. Активность КК-ММ увеличивается также после физической нагрузки, внутримышечных инъекций, хирургических операций. При мышечной дистрофии типа Дюшенна (наследственном, связанным с полом рецессивном заболевании) активность КК-ММ увеличивается в 8-90 раз по сравнению с нормой, активность изофермента увеличена также у носителей этого гена.
Активность КК-ВВ в крови отмечали во время аортокоронарного шунтирования. Полагают, что активность КК-ВВ может быть тестом аноксии тканей. Активность КК-ВВ в крови может также быть следствием гипоксического повреждения мозга, особенно в условиях перинатальной гипоксии. Активность КК-ВВ увеличена у 53% новорожденных с асфиксией. КК-ВВ присутствует в гладкой мускулатуре, но не определяется в сыворотке крови лиц с доброкачественными заболеваниями этих тканей. Одним из возможных объяснений повышения активности КК-ВВ в крови при сосудистых операциях служит предположение, что стенки вен, как, впрочем, и аорты, содержат только одну изоформу КК, а именно КК-ВВ. Активность изофермента КК-ВВ может быть повышена в крови при раке предстательной железы, мелкоклеточном раке легкого, аденокарциноме желудка, лейкозах, хронической почечной недостаточности, передозировке миорелаксантов. Метастазирование рака предстательной железы сопровождается особенно высокими цифрами активности КК-ВВ в крови. Исследователи сходятся во мнении, что активность КК-ВВ может быть использована как неспецифический маркер опухолевого процесса.
В ряде случаев при инфаркте миокарда, а иногда и в его отсутствие высокая активность КК сохраняется неопределенно долго. Наблюдаемые явления обусловлены циркуляцией в крови необычных форм КК и ее изоферментов. После разделения приЭФ фракций КК на электрофореграмме становятся видимыми полосы, не соответствующие положению полос КК-ММ, КК-МВ и КК-ВВ, и можно отметить наличие изоформ КК, движущихся к катоду, что из белков сыворотки крови свойственно иммуноглобулинам. Известно, что окисление сульфгидрильных групп КК изменяет электрофоретическую подвижность молекул, однако при этом они по-прежнему движутся к аноду. У больных метастазирующим раком предстательной железы выделен изофермент КК, мигрирующий между КК-МВ и КК-ВВ.
3.6. Лактатдегидрогеназа(ЛДГ)
Лактатдегидрогеназа (ЛДГ; L-лактат-НАД-оксидоредуктаза, КФ 1.1.1.27) — цинксодержаший фермент, катализирует обратимую реакцию восстановления пировиноградной кислоты в молочную кислоту при участии НАД-Н2.
Фермент в кристаллической форме был получен из мышцы сердца. Подобным же образом были получены кристаллические ферментные препараты из скелетных мышц и печени. При рН 7,0 равновесие реакции смещено в сторону образования лактата, в щелочной среде реакция проходит в обратном направлении. ЛДГ может также реагировать с НАДФ в качестве кофермента, но значительно медленнее, чем с НАД.
ЛДГ — тетрамер; два локуса генов кодируют синтез двух олигомеров — М- и Н-субъединиц. М-субъединица синтезируется главным образом в тканях с анаэробным метаболизмом, в то время как Н-субъединица присутствует в тканях с преобладанием аэробных процессов. Молекулярная масса каждой субъединицы составляет 35 кДа, каждого тетрамера — 140 кДа. Полипептидная цепь обеих субъединиц содержит 330 аминокислотных остатков; различия в их последовательности в субъединицах обнаружены на протяжении более чем 25% длины полипептидной цепи. В тетрамерной структуре ЛДГ субъединицы связаны силами ионных и водородных взаимодействий. На каждой субъединице расположен каталитический центр; диссоциация тетрамера на димеры или мономеры приводит к потере ферментативной активности.
В цитоплазме клеток и сыворотке крови ЛДГ имеет 5 изоферментов, обозначаемых в соответствии с их подвижностью к аноду в электрическом поле: ЛДГ1 (НННН), ЛДГ2 (НННМ), ЛДГ3, (ННММ), ЛДГ4 (НМММ) и ЛДГ5 (ММММ). ЛДГ, участвует в окислении лактата в пируват в тканях с аэробным типом метаболизма (миокард, мозг, почки, эритроциты, тромбоциты). ЛДГ5 оптимизирована природой для превращения пирувата в лактат в тканях с высоким уровнем гликолиза (скелетные мышцы, печень). Не все изоферменты ЛДГ гомогенны: при электрофоретическом разделении изоферментов ЛДГ сыворотки и эритроцитов в полиакриламидном геле обнаружено расщепление ЛДГ3 на две полосы, которое позволяет предположить синтез двух форм ЛДГ3 в тканях. Наличие молекулярных структур двух форм, подтвержденное в реакции с антисывороткой, объясняют различием пространственного (цис и транс) расположения Н-субъединиц и М-субъединиц в тетрамере.
В тканях человека активность ЛДГ на 1 г сухой массы уменьшается в последовательности: почки — скелетная мышца — поджелудочная железа — селезенка — печень — плацента. Изоферменты ЛДГ, и ЛДГ2 преобладают в эритроцитах, лейкоцитах, миокарде, почках, ЛДГ4 и ЛДГ5 — в печени, скелетных мышцах, неопластических тканях, наиболее высокое содержание ЛДГ3 отмечают в лимфоидной ткани, тромбоцитах и опухолях.
Свойства изоферментов ЛДГ определены особенностями входящих в них субъединиц. Изоферментам ЛДГ присущи разные кинетические характеристики: рН, при котором они проявляют максимальную активность, сродство к субстратам и кофакторам.
Клиническое значение определения активности ЛДГ
Активность ЛДГ в сыворотке крови повышается при многих патологических состояниях, поэтому для дифференциальной диагностики заболеваний более целесообразно исследовать изменения спектра изоферментов ЛДГ. В настоящее время накоплено большое количество данных о распределении изоферментов ЛДГ в тканях и об изменении спектра изоферментов ЛДГ в сыворотке крови при различных заболеваниях.
оферментный спектр скелетной мускулатуры показывает преобладание ЛДГ5. При мышечной дистрофии отмечены увеличение более подвижных изоферментов ЛДГ и снижение активности ЛДГ5, что характерно и для многих нейромышечных заболеваний. Причиной изменения спектра изоферментов может быть быстрое удаление малоподвижных изоферментов из циркуляции. Активность ЛДГ5 в сыворотке крови — чувствительный индикатор гепатоцеллюлярного поражения, увеличение его активности обычно наблюдают при гепатите, гипоксии печени (включая застой крови в печени вследствие сердечной недостаточности), лекарственной интоксикации, циррозе, опухолях и травме. Активность ЛДГ в сыворотке крови не повышается при хронических заболеваниях почек и уремии, но иногда возрастает после гемодиализа или плазмафереза, что может быть объяснено удалением из крови ингибиторов (мочевина, оксалаты).
Общая активность ЛДГ при инфаркте миокарда наиболее значительно повышается в течение первых 2 сут после приступа стенокардии и до исходного уровня понижается медленно, в течение 14-16 дней, эпизодическое повышение ЛДГ можно отметить и в более поздние сроки.
Активность ЛДГ подвержена гормональному влиянию. Большие дозы тироксина снижали синтез фермента, при этом в большей мере отмечено ингибирование синтеза субъединицы М. Норадреналин и адреналин вызывают увеличение общей активности ЛДГ с преобладанием активности ЛДГ1, и ЛДГ2. Активность фермента в крови возрастает при действии анаболических стероидов и этанола, а также ряда медикаментозных препаратов — клофибрата, кофеина, сульфаниламидов и др.
Спектр изоферментов ЛДГ может меняться при неопластических процессах. В таких случаях его трудно интерпретировать, так как источником изоферментов ЛДГ служит не только неопластическая ткань, но и ткани, разрушаемые метастазами. Однако изоферментный спектр транссудатов при опухолевом поражении сходен с таковым сыворотки крови, тогда как в воспалительных экссудатах преобладает активность ЛДГ1 и ЛДГ2. Для ЛДГ, как и для других ферментов, при опухолевом процессе характерен синтез изоферментов, свойственных эмбриональным тканям. Недифференцированные эмбриональные ткани имеют спектр изоферментов ЛДГ, в котором преобладают ЛДГ2 и ЛДГ3, а также ЛДГ4. В злокачественных опухолях обнаружено три вида распределения изоферментов ЛДГ. Увеличение содержания ЛДГ4 и ЛДГ5 выявлено при опухолях предстательной железы, матки, молочных желез, желудка, толстой кишки, мочевого пузыря и некоторых типах опухолей мозга. У больных лейкозом, злокачественной лимфомой, нейробластомой, феохромоцитомой, а также при опухолях полости рта, раке бронхов и некоторых типах опухолей мозга увеличена активность ЛДГ2, ЛДГ3, ЛДГ4. Увеличение активности ЛДГ, отмечено в сыворотке крови больных с некоторыми типами опухолей мозга и разными типами опухолей половых органов.
Возможность повышения активности ЛДГ, при опухолевом процессе следует учитывать при диагностике инфаркта миокарда. Иногда при опухолях мозга, раке пишевода, нейробластоме отмечают необычную дополнительную полосу приЭФ сыворотки крови и ткани опухоли. Определение спектра изоферментов ЛДГ в сыворотке крови при онкологических заболеваниях полезно не только для диагностики, но и для контроля эффективности лечения. Обнаружено, что нормализация спектра изоферментов ЛДГ коррелирует с успешностью ответа больного на лечение.
Наличие осложнений при инфаркте миокарда и сопутствующие заболевания могут изменить спектр ЛДГ и активность ЛДГ. Выявление спектра изоферментов, характерного для инфаркта миокарда, возможно при застое крови в печени и почках вследствие сердечной недостаточности, при ишемическом поражении некоторых органов из-за резкого снижения сердечного выброса. При эмболии легочной артерии, которую в ряде случаев приходится дифференцировать с инфарктом миокарда, увеличение в крови активности ЛДГ2 и ЛДГ3 может быть объяснено выходом ферментов из тромбоцитов, патологией печени, вызванной венозной гипертензией, анемией коркового слоя надпочечников и почек. Поскольку эти нарушения не всегда удается различить, изменение спектра изоферментов ЛДГ интерпретировать непросто.
В сыворотке крови тяжелобольных (как правило, в терминальном состоянии) методом ЭФ на ацетате целлюлозы выявляется дополнительная полоса, более близкая к катоду, чем ЛДГ5, названная ЛДГ6. Во всех наблюдениях в тканях печени, скелетных мышцах, почке, селезенке и надпочечниках также отмечено присутствие ЛДГ6; в ряде случаев появление этой фракции носило транзиторный характер, ЛДГ6 не обнаружена в тканях миокарда. Считают, что нет нозологической формы заболевания, для которой характерна ЛДГ6, но есть тяжелые клинические состояния, обусловливающие ее появление, к которым следует отнести выраженный ацидоз, гипотонию и сепсис.
Источник: studopedia.su
Структурно-функциональная организация ферментов
В составе фермента выделяют области, выполняющие различную функцию:
1. Активный центр – комбинация аминокислотных остатков (обычно 12-16), обеспечивающая непосредственное связывание с молекулой субстрата и осуществляющая катализ. Аминокислотные радикалы в активном центре могут находиться в любом сочетании, при этом рядом располагаются аминокислоты, значительно удаленные друг от друга в линейной цепи. В активном центре выделяют два участка:
- якорный (контактный, связывающий) – отвечает за связывание и ориентацию субстрата в активном центре,
- каталитический – непосредственно отвечает за осуществление реакции.
Схема строения ферментов
У ферментов, имеющих в своем составе несколько мономеров, может быть несколько активных центров по числу субъединиц. Также две и более субъединицы могут формировать один активный центр.
У сложных ферментов в активном центре обязательно расположены функциональные группы кофактора.
Схема формирования сложного фермента
2. Аллостерический центр (allos – чужой) – центр регуляции активности фермента, который пространственно отделен от активного центра и имеется не у всех ферментов. Связывание с аллостерическим центром какой-либо молекулы, называемой активатором или ингибитором (или эффектором, модулятором, регулятором), вызывает изменение конфигурации белка-фермента и, как следствие, скорости ферментативной реакции.
Аллостерические ферменты являются полимерными белками, активный и регуляторный центры находятся в разных субъединицах.
Схема строения аллостерического фермента
В качестве такого регулятора может выступать продукт данной или одной из последующих реакций, субстрат реакции или иное вещество (см «Регуляция активности ферментов»).
Изоферменты
Изоферменты – это молекулярные формы одного и того же фермента, возникшие в результате небольших генетических различий в первичной структуре фермента, но катализирующие одну и ту же реакцию. Изоферменты отличаются сродством к субстрату, максимальной скоростью катализируемой реакции, чувствительностью к ингибиторам и активаторам, условиями работы (оптимум pH и температуры).
Как правило, изоферменты имеют четвертичную структуру, т.е. состоят из двух или более субъединиц. Например, димерный фермент креатинкиназа (КК) представлен тремя изоферментными формами, составленными из двух типов субъединиц: M (англ. muscle – мышца) и B (англ. brain – мозг). Креатинкиназа-1 (КК-1) состоит из субъединиц типа B и локализуется в головном мозге, креатинкиназа-2 (КК-2) – по одной М- и В-субъединице, активна в миокарде, креатинкиназа-3 (КК-3) содержит две М-субъединицы, специфична для скелетной мышцы. Определение активности разных изоферментов КК в сыворотке крови имеет клинико-диагностическое значение.
Изоферменты креатинкиназы |
Изоферменты лактатдегидрогеназы |
Также существует пять изоферментов лактатдегидрогеназы (роль ЛДГ) – фермента, участвующего в обмене глюкозы. Отличия между ними заключаются в разном соотношении субъединиц Н (англ. heart – сердце) и М (англ. muscle – мышца). Лактатдегидрогеназы типов 1 (Н4) и 2 (H3M1) присутствуют в тканях с аэробным обменом (миокард, мозг, корковый слой почек), обладают высоким сродством к молочной кислоте (лактату) и превращают его в пируват. Изоферменты ЛДГ-4 (H1M3) и ЛДГ-5 (М4) находятся в тканях, склонных к анаэробному обмену (печень, скелетные мышцы, кожа, мозговой слой почек), обладают низким сродством к лактату и катализируют превращение пирувата в лактат. В тканях с промежуточным типом обмена (селезенка, поджелудочная железа, надпочечники, лимфатические узлы) преобладает ЛДГ-3 (H2M2). Определение активности разных изоферментов ЛДГ в сыворотке крови имеет клинико-диагностическое значение.
Еще одним примером изоферментов является группа гексокиназ, которые присоединяют фосфатную группу к моносахаридам гексозам и вовлекают их в реакции клеточного метаболизма. Из четырех изоферментов выделяется гексокиназа IV (глюкокиназа), которая отличается от остальных изоферментов высокой специфичностью к глюкозе, низким сродством к ней и нечувствительностью к ингибированию продуктом реакции.
Мультиферментные комплексы
В мультиферментном комплексе несколько ферментов прочно связаны между собой в единый комплекс и осуществляют ряд последовательных реакций, в которых продукт реакции непосредственно передается на следующий фермент и является только его субстратом. Возникает туннельный эффект, т.е. субстрат попадает в созданный ферментами «туннель». В результате промежуточные метаболиты избегают контакта с окружающей средой, снижается время их перехода к следующему активному центру и значительно ускоряется скорость реакции.
Строение мульферментного комплекса
Например,
- пируватдегидрогеназный комплекс (пируватдегидрогеназа), превращающий пируват в ацетил-SКоА,
- α-кетоглутаратдегидрогеназный комплекс (в цикле трикарбоновых кислот) превращающий α-кетоглутарат в сукцинил-SКоА,
- комплекс под названием «синтаза жирных кислот» (или пальмитатсинтаза), синтезирующий пальмитиновую кислоту
- карбамоилфосфатсинтетаза, первый фермент синтеза мочевины в печени и синтеза пиримидиновых оснований
Абзимы
Абзимами называются антитела, имеющие каталитическую функцию (англ. abzymes, antibodies as enzymes) и катализирующие конкретные реакции. Такая способность возникает в результате формирования промежуточного продукта при связывании антитела с антигеном (имитация переходного комплекса E-X ферментативной реакции).
Источник: biokhimija.ru
Диагностическое значение изоферментов
Исследование активности изоферментов КФК в сыворотке крови имеет важное диагностическое значение. Изоферменты креатинкиназы особо важно исследовать при остром инфаркте миокарда, так как МВ-форма в значительном количестве содержится практически только в сердечной мышце. Повышение активности МВ-формы в сыворотке крови свидетельствует о поражении именно сердечной мышцы.
Возрастание активности ферментов сыворотки крови при многих патологических процессах объясняется, прежде всего, двумя причинами:
- выходом в кровяное русло ферментов из поврежденных участков органов или тканей на фоне продолжающегося их биосинтеза в поврежденных тканях;
- одновременным повышением каталитической активности некоторых ферментов, переходящих в кровь. Возможно, что повышение активности ферментов при «поломке» механизмов внутриклеточной регуляции обмена веществ связано с прекращением действия соответствующих регуляторов и ингибиторов ферментов, изменением под влиянием различных факторов строения и структуры макромолекул ферментов.
Содержание фракции ММ изоферментов повышается:
- при травмах скелетных мышц,
- повреждении сердечной мышцы или ткани мозга;
- при таких заболеваниях мышц,
- как дистрофия,
- дерматомиозит,
- полимиозит,
- при пониженной функции щитовидной железы;
- при рабдомиолизе,
- после тяжелой физической нагрузки.
Повышенное содержание изофермента МВ отмечается сразу после инфаркта миокарда (через 2-4 часа) и сохраняется в течение примерно 3 суток. Если по истечении этого срока уровень изофермента МВ не возвращается к норме, то это может свидетельствовать о распространении инфаркта или о новом инфаркте. Содержание изофермента МВ тоже наблюдается при рабдомиолизе, повреждении мышц, а также синдроме Рея.
Повышение уровня изофермента ВВ иногда происходит при тяжелом шоке, атрезии желчных протоков, а также при некоторых злокачественных опухолях (карциноме яичников, молочной железы простаты).
Методы определения активности КФК
Унифицированным методом определения активности креатинфосфокиназы служит метод с использованием в качестве субстрата реакции креатина. Активность фермента пропорциональна количеству неорганического фосфора, образующегося в результате кислотного гидролиза продукта креатинкиназной реакции – креатинфосфата.
Неорганический фосфор определяется по цветной реакции с молибдатом аммония. В норме активность креатинкиназы в крови, установленная с помощью этого метода, достигает 100 нмоль/(с*л), или до 6 МЕ. Активность МВ-формы креатинкиназы составляет менее 2 % от общей активности фермента.
Вторым методом определения активности креатинкиназы, также принятым в качестве унифицированного, является метод с использованием в качестве субстрата креатинфосфата. Активность фермента пропорциональна количеству креатина, образующегося в результате ферментативной реакции. Креатин определяется по цветной реакции с a-нафтолом.
В норме активность креатинкиназы в сыворотке крови, установленная с помощью этого метода, составляет от 0 до 220 нмоль/(с*л), или от 0 до 13 МЕ [Международная единица = 16,67 нмоль/(с*л)].
Видео: АЛТ и АСТ в биохимическом анализе крови
Источник: MedAnalises.net
Изоферменты – это множественные формы одного фермента, катализирующие одну и ту же реакцию, но отличающие по физическим и химическим свойствам (сродству к субстрату, максимальной скорости катализируемой реакции, электрофоретической подвижности, разной чувствительности к ингибиторам и активаторам, оптимуму рН и термостабильности). Изоферменты имеют четвертичную структуру, которая образована четным количеством субъединиц (2, 4, 6 и т.д.). Изоформы фермента образуются в результате различных комбинаций субъединиц.
В качестве примера можно рассмотреть лактатдегидрогеназу (ЛДГ), фермент, который катализирует обратимую реакцию:
НАДН2 НАД+
пируват ←ЛДГ→ лактат
ЛДГ существует в виде 5 изоформ, каждая из которых состоит из 4-х протомеров (субъединиц) 2 типов М (muscle) и Н (heart). Синтез протомеров М и Н типа кодируется двумя разными генетическими локусами. Изоферменты ЛДГ различаются на уровне четвертичной структуры: ЛДГ1(НННН), ЛДГ2(НННМ), ЛДГ3(ННММ), ЛДГ4(НМММ), ЛДГ5(ММММ).
Полипептидные цепи Н и М типа имеют одинаковую молекулярную массу, но в составе первых преобладают карбоновые аминокислоты, последних – диаминокислоты, поэтому они несут разный заряд и могут быть разделены методом электрофореза.
Кислородный обмен в тканях влияет на изоферментный состав ЛДГ. Где доминирует аэробный обмен, там преобладают ЛДГ1, ЛДГ2(миокард, надпочечники), где анаэробный обмен — ЛДГ4, ЛДГ5(скелетная мускулатура, печень). В процессе индивидуального развития организма в тканях происходит изменение содержания кислорода и изоформ ЛДГ. У зародыша преобладают ЛДГ4, ЛДГ5. После рождения в некоторых тканях происходит увеличение содержания ЛДГ1, ЛДГ2.
Существование изоформ повышает адаптационную возможность тканей, органов, организма в целом к меняющимся условиям. По изменению изоферментного состава оценивают метаболическое состояние органов и тканей.
Изоформы креатинкиназы.Креатинкиназа (КК) катализирует реакцию образования креатинфосфата:
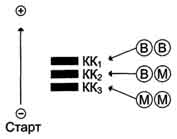
Молекула КК — димер, состоящий из субъединиц двух типов: М (от англ, muscle — мышца) и В (от англ, brain — мозг). Из этих субъединиц образуются 3 изофермента — ВВ, MB, MM. Изофермент ВВ находится преимущественно в головном мозге, ММ — в скелетных мышцах и MB — в сердечной мышце. Изоформы КК имеют разную электрофоретическую подвижность (рис. 2-36).
Активность КК в норме не должна превышать 90 МЕ/л. Определение активности КК в плазме крови имеет диагностическое значение при инфаркте миокарда (происходит повышение уровня МВ-изоформы). Количество изоформы ММ может повышаться при травмах и повреждениях скелетных мышц. Изоформа ВВ не может проникнуть через гематоэнцефалический барьер, поэтому в крови практически не определяется даже при инсультах и диагностического значения не имеет.
Источник: studopedia.ru
Источник: www.chem21.info
